 2022, Vol. 39
2022, Vol. 39



2. 新疆土壤与植物生态过程重点实验室,乌鲁木齐 830052
2. Key Laboratory of Soil and Plant Ecological Process, Urumqi 830052, China
新疆巴音布鲁克高寒草原是我国第一大亚高山高寒草原,也是新疆主要的优质放牧草地之一[1]。由于自然条件和人为活动干扰,巴音布鲁克草原出现草地退化、土壤沙化、害草蔓延等一系列生态问题[2],其中放牧压力是导致草地退化的主要原因之一[3]。放牧会引起土壤体积密度、含水量和温度等非生物性质的变化,更会通过对地上植物的影响进而改变土壤动物群落特征[4]。土壤线虫是土壤中数量和多样性最为丰富的微型土壤动物之一,也是草地生态系统碎屑食物网的优势类群,其食性多样,在土壤食物网中占据多个营养级地位,对草地土壤生态系统的物质循环、能量流动及地上植物群落的组成、演替及生产力具有重要的调控作用[5-7]。土壤线虫因其营养类群多样,对自然及人为扰动响应敏感,被看作评价草地土壤生态系统健康状况的典型指示生物[8-9]。在放牧对草地生态系统土壤线虫的影响方面,大量研究认为放牧强度对土壤线虫群落的营养类群有显著影响,放牧可通过牲畜采食践踏等活动改变植物群落,进而影响食细菌类线虫和植食类线虫的数量[10-11]。内蒙古半干旱草原土壤动物群落的多样性、均匀性和种类丰富度会随放牧强度增加而降低,放牧强度增加使土壤线虫食物网退化、不稳定[12]。为探讨新疆高寒草地土壤线虫群落对放牧强度的响应,本研究对巴音布鲁克高寒草地三个长期放牧强度的样地土壤线虫群落结构及环境因素进行检测分析,了解放牧强度干扰对高寒草地土壤线虫群落结构特征的影响,评价放牧强度对高寒草地土壤生态系统健康的干扰性,为合理利用巴音布鲁克高寒草地资源以及维持草地生态系统健康提供理论依据。
1 材料与方法 1.1 研究区概况研究区位于新疆维吾尔自治区巴音郭楞蒙古自治州和静县境内(82°59′ ~84°35′ E,42°40′ ~43°00′ N),海拔2 300~3 042 m,年均降水量273 mm,年平均气温-4.6 ℃,属于典型的高寒气候。依据载畜量确定三个长期不同放牧强度的样地,载畜量(只·a-1·hm-2,以羊数量计)=暖季载畜量+冷季载畜量=暖季放牧数量/牧食面积×暖季放牧时间/365+冷季放牧数量/牧食面积×冷季放牧时间/365[13]。放牧强度(以羊数量计)分别为轻度放牧(0.65只·hm-2)、中度放牧(2.09只· hm-2)、重度放牧(4.15只·hm-2)[14]。研究区各样地基本情况见表 1。
|
|
表 1 巴音布鲁克高寒草地各样地基本情况 Table 1 Basic conditions of alpine grassland in Bayanbulak |
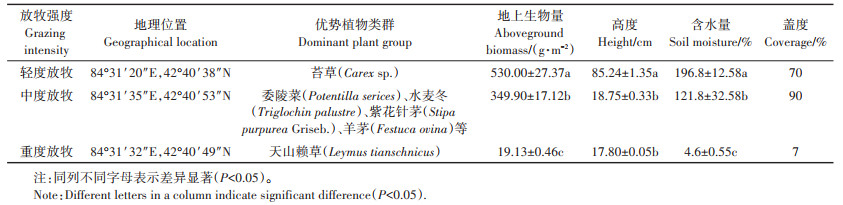
在三种放牧强度样地内各设置1条100 m长的样线,在每条样线上以20 m为间隔布设一个1 m×1 m样方,即每条样线有5个重复样方,共15个样方。在样方内进行植被分种、数量和盖度调查,样方内植物分物种将地面以上所有绿色部分用剪刀齐地面剪下后,按样方编号和物种分类,分别装进信封袋,作好标记,称取地上植被生物量(鲜质量),具体情况见表 1。每个样方采集0~5、5~10、10~15、15~20 cm和20~25 cm土壤样品并监测土壤含水量。将每个采样单元内各个采样点的土壤混合均匀后,按四分法获取足量的样品,放入无菌自封袋中。采集的土样1份冰袋冷藏保存用于分离土壤线虫,1份自然风干保存用于基本理化性质测定。
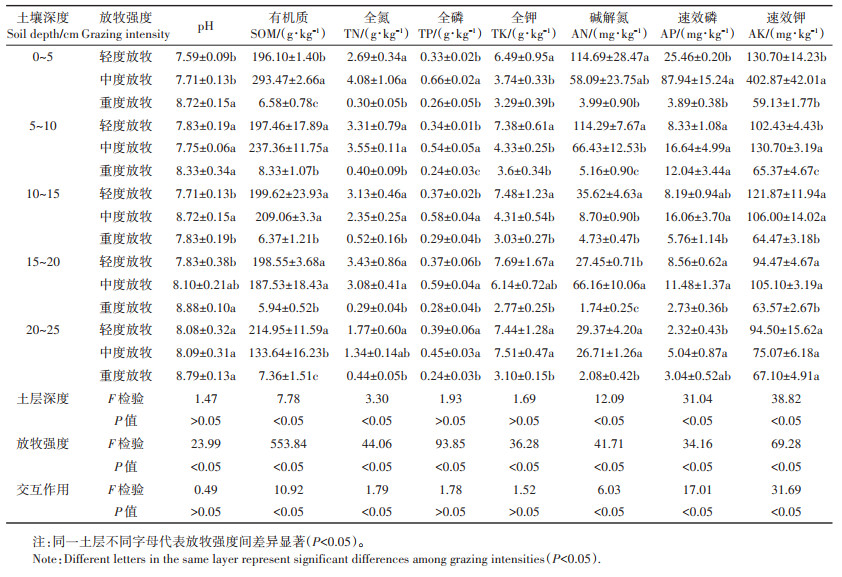
1.3 土壤环境因子测定土壤养分测定参考《土壤农业化学分析方法》[15],有机质(SOM)测定采用重铬酸钾外加热法;全磷(TP)测定采用磷钼蓝比色法;全钾(TK)测定采用火焰光度计法;全氮(TN)测定采用高氯酸-硫酸消化法;速效磷(AP)测定采用碳酸氢钠-硫酸钼锑抗比色法;速效钾(AK)测定采用乙酸铵提取法;碱解氮(AN)测定采用扩散法。土壤pH采用pH计(PHSJ- 4F,上海仪电科学仪器股份有限公司)测定(土水比1∶5),土壤含水量(SWC)采用烘干法进行测定。
1.4 土壤线虫群落分析称取不同放牧强度下土壤鲜样50 g,采用改进的浅盘分离-蔗糖离心浮选的连续提取法进行线虫分离[16-18],分离出的线虫数量用解剖镜计数,并折算成每100克干土中线虫的数量,每个样品随机选择200条线虫进行制片(少于200条的全部制片),采用光学显微镜鉴定,线虫鉴定参照文献[19-20],根据线虫的取食习性和生活史特征将线虫划分为4种营养类群:食细菌类(Bacterivores)、食真菌类(Fungivores)、植物寄生类(Plant-parasites)、捕食/杂食类(Predators/Omnivores),并计算土壤线虫群落生态指数[21]。

|
(1) |

|
(2) |
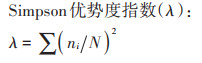
|
(3) |

|
(4) |

|
(5) |

|
(6) |
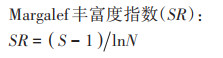
|
(7) |
式中:ni为第i类群的个体数;N为群落所有类群的个体总数;NB为食细菌类线虫数量;NF为食真菌类线虫数量;NP为植物寄生类线虫总数;pi为第i个分类单元中个体占自由生活线虫个体数量的比例;S为鉴定分类单元的数目;Vi为根据自由生活线虫在生态演替中的不同生活策略分别赋予的c-p值(1~5);fi为植物寄生类土壤线虫第i个分类单元中个体数量。
1.5 数据处理土壤线虫数量折算成每100 g干土含有线虫的条数,丰度以不同属或营养类群的线虫数量占线虫总数的百分比(%)来表示[22],采用Excel 2016软件进行数据统计与处理,用SPSS 24.0软件对土壤pH、线虫总数、各营养类群丰度以及生态学指数进行单因素方差分析(One-way ANOVA)和双因素方差分析(Two-way ANOVA),使用Origin 2018绘图,并使用CANOCO 5.0对土壤线虫群落结构和土壤环境因子的相关性(Redundancy analysis,RDA)进行排序。
2 结果与分析 2.1 放牧强度对土壤环境因子的影响不同放牧强度下,随着土层深度的增加土壤各养分含量总体呈下降趋势。在中度放牧下土壤有机质、全氮、全磷、速效磷和速效钾显著高于轻度放牧和重度放牧(P < 0.05);轻度放牧的全钾和碱解氮显著高于中度放牧和重度放牧(P < 0.05),重度放牧的土壤养分指标均低于轻度放牧和中度放牧。重度放牧土壤pH平均值最高,轻度放牧土壤pH平均值最低。双因素方差分析表明,放牧强度、土层深度及其两者的交互作用对土壤有机质、全氮、碱解氮、速效磷和速效钾均具有显著影响(P < 0.05),放牧强度对pH、全磷和全钾具有显著影响,而土层深度和两者交互作用对pH、全磷和全钾无显著影响(表 2)。
|
|
表 2 放牧强度对土壤环境因子的影响(均值±标准误) Table 2 The influence of grazing intensity on soil environmental factors(Mean±standard error) |
土壤线虫数量在不同放牧强度土壤间存在显著差异(P < 0.05),如图 1所示,土壤线虫主要分布在0~5 cm土层中,随土层深度的增加,土壤线虫的数量逐渐减少。在中度放牧下,0~5 cm土层线虫个体密度显著高于轻度放牧和重度放牧(P < 0.05),而轻度放牧和重度放牧之间无显著差异。

|
图 1 土壤线虫个体密度的垂直分布 Figure 1 Vertical distribution of individual density of soil nematodes |
三种放牧强度样地共获得土壤线虫1 698条,线虫个体密度为每100 g干土93条,分属于32科38属,轻度放牧和中度放牧的共同优势属为绕线属(Plectus),前矛线属(Prodorylamus),与重度放牧无共同优势属(表 3)。
|
|
表 3 土壤线虫群落组成及营养类群c-p值 Table 3 Composition of soil nematode community and c-p value of nutrient group |
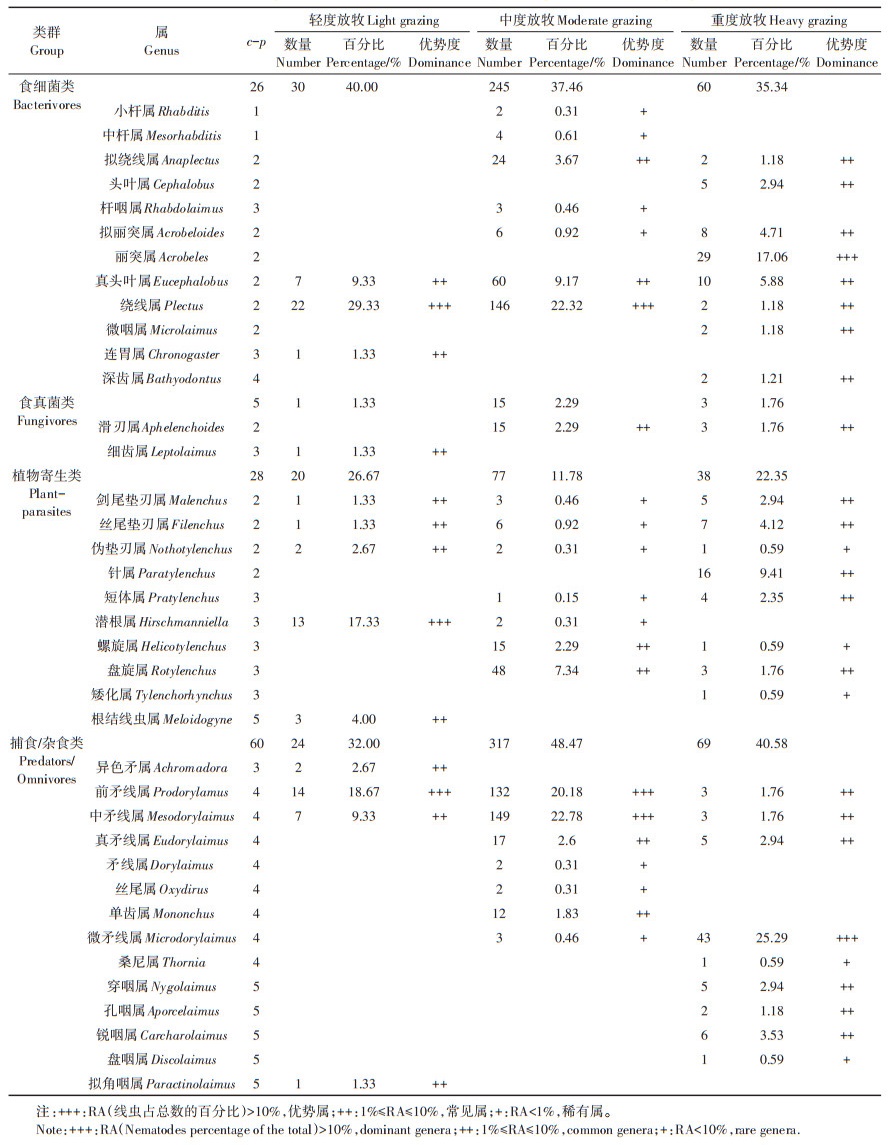
轻度放牧0~25 cm土层共分离得到土壤线虫80条,个体密度为每100 g干土17条,鉴定线虫75条,分属于13属,其中绕线属、潜根属(Hirschmanniella)、前矛线属为优势属,分别占群落中线虫个体总数的29.33%、17.33%、18.67%。真头叶属(Eucephalobus)等10属为常见属,占所有已知个体总数的34.67%;未发现稀有属。
中度放牧0~25 cm土层共分离得到土壤线虫1 427条,个体密度为每100 g干土249条,鉴定线虫654条,分属于22属。其中绕线属、前矛线属、中矛线属(Mesodorylaimus)为优势属,分别占群落中线虫个体总数的22.32%、20.18%、22.78%;拟绕线属(Anaplectus)等7属为常见属,占所有已知个体数的29.19%;小杆(Rhabditis)等12属为稀有属,占所有已知个体总数的5.53%。
重度放牧0~25 cm土层共分离得到土壤线虫191条,个体密度为每100 g干土14条,鉴定线虫170条,分属于26属。其中丽突属(Acrobeles)和微矛线属(Microdorylaimus)为优势属,分别占群落中线虫个体总数的17.06%、25.29%;拟绕线属(Anaplectus)等19属为常见属,占所有已知个体数的54.70%;伪垫刃(Nothotylenchus)等5属为稀有属,占所有已知个体总数的2.95%。
综上得出不同放牧强度会影响巴音布鲁克高寒草地土壤线虫群落结构特征,放牧强度的增加会导致土壤线虫个体密度减少、丰富度增加。不同放牧强度下,土壤线虫的组成存在一定的差异。
从图 2三种放牧强度下线虫营养类群分布可以看出,随着土层深度的增加,土壤线虫各营养类群所占比例发生一定的变化。在轻度放牧强度下,0~5 cm土层的捕食/杂食类线虫占比显著高于其他土层,20~ 25 cm土层的植物寄生类线虫占比显著高于其他土层;中度放牧样地5~10 cm土层中食细菌类线虫、食真菌类线虫和植物寄生类线虫的总占比显著高于0~5 cm土层;重度放牧样地捕食/杂食类线虫占比随土层深度的增加呈降低的趋势。

|
图 2 土壤线虫群落不同营养类群空间分布特征 Figure 2 Spatiotemporal distribution of different nutrient groups in soil nematode community |
多样性指数(H)、均匀度指数(J)、优势度指数(λ)及丰富度指数(SR)可以用来表征巴音布鲁克高寒草地土壤线虫群落的生物多样性(表 4)。比较三种放牧强度下不同土层的H值可知,轻度放牧下土壤线虫的H值最低,平均为0.95;重度放牧下土壤线虫的J值和SR值最高,平均值分别为0.91和2.27;轻度放牧下线虫的H值和λ值显著低于中度放牧和重度放牧(P < 0.05),各放牧强度的线虫J值无显著差异;土壤线虫群落的SR值随着放牧强度的增大而增加,轻度放牧与重度放牧下线虫的SR值具有显著差异(P < 0.05)。
|
|
表 4 土壤线虫群落生态指数 Table 4 Ecological index of soil nematode community |
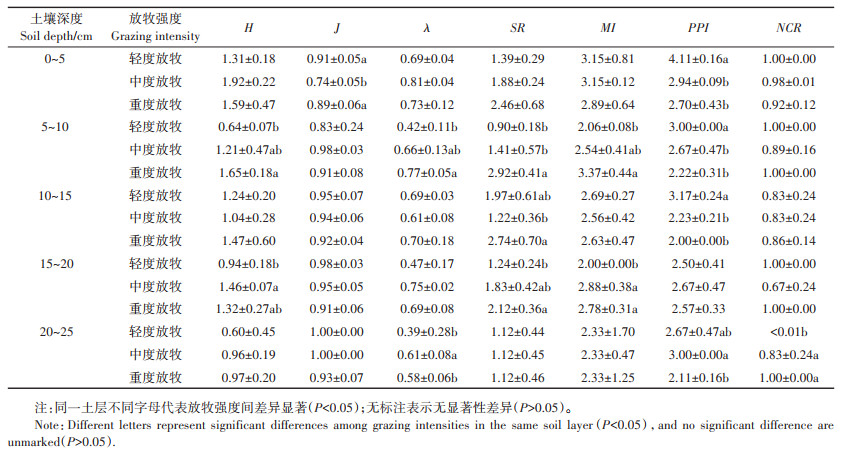
土壤线虫群落的功能结构特征可用自由生活线虫成熟度指数(MI)、植物寄生类线虫成熟度指数(PPI)、线虫通道指数(NCR)来表征。三种放牧强度下不同土层的MI值介于2.00~3.37之间,平均值为2.67。轻度放牧5~10 cm土层MI值显著低于重度放牧,在15~20 cm土层显著低于中度放牧和重度放牧(P < 0.05)。PPI值介于2.00~4.11之间,平均值为2.70。0~15 cm土层轻度放牧PPI值显著高于中度放牧(P < 0.05),重度放牧与中度放牧间无显著差异。NCR值介于0~1.00之间,平均值为0.85,说明该土壤中有机物分解方式以细菌分解为主。
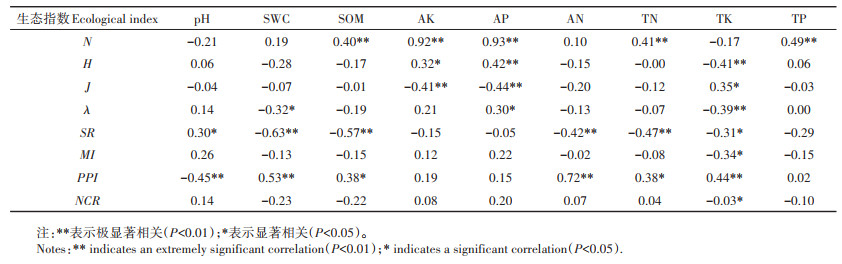
2.5 土壤线虫群落特征与土壤环境因子的关系由图 3可知,不同放牧强度下,各土壤环境因子对土壤线虫群落的影响不一致。轻度放牧强度下土壤速效磷、含水量和全钾对土壤线虫群落的影响最大;中度放牧强度下土壤全氮、速效磷和速效钾对土壤线虫群落的影响最大;重度放牧强度下土壤含水量、全磷和速效磷对土壤线虫群落的影响最大。轻度放牧和中度放牧强度下土壤线虫前矛线属(Prodorylamus)与土壤含水量具有较强的正相关性,真头叶属(Eucephalobus)在中度放牧和重度放牧强度下都与全磷有较好的正相关性。为了能够综合反映土壤线虫群落特征指数与土壤环境因子的关系,对二者进行相关性分析(表 5)。土壤环境因子对土壤线虫群落特征指数的影响不同,速效磷、速效钾和pH是影响土壤线虫群落多样性的重要指标,具有显著相关性;有机质、碱解氮、速效磷、速效钾和全磷极显著影响土壤线虫的数量;土壤含水量和全钾极显著影响土壤线虫的功能结构。

|
图 3 不同放牧强度下土壤线虫群落与土壤环境因子间的冗余分析(RDA) Figure 3 RDA of soil nematode community and soil environmental factors under different grazing intensities |
|
|
表 5 土壤线虫群落特征指数与土壤环境因子的相关性分析(r) Table 5 Correlation analysis between soil nematode community characteristic index and soil environmental factors(r) |
土壤线虫群落结构和多样性变化的影响因素包括土壤因素、气候因素、植物因素、动物因素和微生物因素等[23]。从巴音布鲁克高寒草地三种放牧强度的样地中共获得土壤线虫1 698条,平均每100 g干土中93条,分属于32科38属,与青藏高原东缘高寒草甸放牧样地相比少4属[24]。前人研究结果表明放牧强度会降低土壤线虫的总数量和优势类群,土壤线虫数量随着土层深度的增加而减少[25-26]。线虫类群属数和数量具有表聚性特征,主要生活在0~5 cm土层[27]。本研究结果同样表现出相似分布特征。本研究发现中度放牧样地的土壤线虫数量及各个营养类群的线虫数量都明显高于轻度放牧样地和重度放牧样地,综合0~25 cm土层中的各指标来看,中度放牧样地的线虫多样性指数最高,线虫数量最多,均匀度指数高,优势度指数低,说明生态系统中土壤线虫多样性较高,随着土层深度的增加,土壤线虫群落生物多样性降低。其原因可能是由于轻度放牧样地处于淹水状态,地上植被单一,植株过高,而重度放牧样地已经严重退化,土壤沙化严重,含水量低,地上植被单一,覆盖度低,但还是有羊群不断踩踏,粪尿进入草地。中度放牧样地的植物群落丰富,覆盖度高,是羊群主要摄食的地方,其土壤养分高,土壤微生物群落丰富,食物来源也更加丰富,导致各个营养类群的土壤线虫数量都明显高于其他样地。
线虫通道指数(NCR)、自由生活线虫成熟度指数(MI)和植物寄生类线虫成熟度指数(PPI)反映土壤线虫类群功能结构特征和生活史的多样性[6],可以用来评价人类干扰活动对土壤线虫群落的影响。MI值越低,表明环境系统所受的干扰越大[28-29]。本研究发现轻度放牧样地MI平均值均低于中度放牧和重度放牧,说明环境系统所受的干扰大,但是该地区受放牧干扰较小,其原因可能是土壤水分含量高,导致线虫的生长发育受到阻碍,线虫数量少,这与薛会英等[30]的研究结果一致。综合三个放牧强度0~25 cm的PPI值来看,重度放牧 < 中度放牧 < 轻度放牧,说明随着放牧强度的增加,地上植被生长状况越来越弱。植物的丰富度和密度通过地下根对植物寄生类线虫产生影响,这种影响比放牧干扰对植物寄生类线虫的影响更加剧烈。NCR值代表微生物参与分解有机质的途径,NCR值为0,代表土壤有机质完全依靠真菌分解;若NCR值为1,则表示有机质完全由细菌分解[31]。三种不同放牧强度下NCR值在0~1之间,平均值为0.85,说明细菌是有机质的主要分解者。细菌占土壤微生物90%以上[32],而细菌的主要食物来源为低碳氮比的有机质,易降解,物质循环速率快,有利于植物从土壤中吸收养分[33]。
不同放牧强度和环境因子影响土壤线虫群落结构特征,草地放牧下土壤养分一般受地面活动影响大,造成土壤上层养分高于深层[34]。本研究结果表明不同放牧强度的干扰下土壤上层养分含量高于深层,由于长期放牧导致土壤退化严重,重度放牧样地的土壤养分显著低于轻度放牧样地和中度放牧样地。中度退化样地的土壤有机质、速效钾、速效磷也都明显高于轻度退化样地和重度退化样地。该研究结果与前人研究结果一致,说明土壤有机物和微生境也是影响土壤线虫的主要因素[30]。研究中土壤速效磷、有机质、速效钾、全氮和全磷与土壤线虫数量呈极显著正相关,其中速效磷在冗余分析中与土壤线虫数量呈现较好的相关性,是主要影响因子,表明速效磷、有机质、速效钾、全氮和全磷对土壤线虫数量有不可忽视的影响,进而影响群落结构。前人研究结果表明地上植物群落和土壤环境因子的改变都会影响土壤线虫群落组成,土壤线虫群落的变化与地上植物具有极强的相关性[7],这与本研究结果一致,说明放牧强度的变化可能会使植物根系生物量和分泌物改变,从而使作为初级消费者的植物寄生类线虫和食微线虫数量发生变化,而捕食/杂食线虫作为次级消费者也随之改变。土壤中各种营养成分的改变会对土壤食物网的复杂程度以及土壤线虫群落结构造成显著影响[7, 35]。本研究仅对巴音布鲁克高寒草地三种放牧强度下土壤线虫群落特征进行了初步研究,但对于不同放牧条件下,土壤线虫群落对环境变化响应机制的研究还需要结合生物和非生物因素进一步开展,这对于从生物学角度评价草地土壤生态系统健康具有重要意义,可为巴音布鲁克高寒草地资源的合理利用以及维持草地生态系统健康提供理论依据。
4 结论(1)巴音布鲁克高寒草地不同放牧强度下土壤线虫数量随土层深度的增加而降低。中度放牧强度(2.09只·hm-2)下土壤线虫数量最多,其中食细菌类线虫、食真菌类线虫、植物寄生类线虫和捕食/杂食类线虫的数量最大。重度放牧强度(4.15只·hm-2)下土壤线虫的种类最多。
(2)巴音布鲁克高寒草地在轻度放牧(0.65只· hm-2)和中度放牧强度下土壤线虫的优势属主要为绕线属和前矛线属;重度放牧强度下土壤线虫的优势属为丽突属和微矛线属。
(3)放牧强度影响土壤线虫群落结构,中度放牧强度下土壤线虫个体密度最高,土壤生态系统食物网相对稳定,有利于高寒草地生态系统的维持。
| [1] |
周瑞莲, 赵哈林. 高寒山区人工草地退化的生理生态学机理[J]. 中国草地, 1997(4): 6-11. ZHOU R L, ZHAO H L. Analysis on physiogical and ecological causes of the degeneration of artificial grassland in the alpine meadew[J]. Grassland of China, 1997(4): 6-11. |
| [2] |
阿依敏·波拉提, 安沙舟, 董乙强, 等. 巴音布鲁克高寒草原不同退化程度土壤化学计量比特征[J]. 新疆农业科学, 2018, 55(5): 957-965. AYIMIN B, AN S Z, DONG Y Q, et al. Soil stoichiometry characteristics in different degradation stages of alpine steppe in Bayanbulak[J]. Xinjiang Agricultural Sciences, 2018, 55(5): 957-965. |
| [3] |
王德利, 王岭, 辛晓平, 等. 退化草地的系统性恢复: 概念、机制与途径[J]. 中国农业科学, 2020, 53(13): 2532-2540. WANG D L, WANG L, XIN X P, et al. Systematic restoration for degraded grasslands: Concept, mechanisms and approaches[J]. Scientia Agricultura Sinica, 2020, 53(13): 2532-2540. DOI:10.3864/j.issn.0578-1752.2020.13.002 |
| [4] |
BRIAR S S, CULMAN S W, YOUNG-MATHEWS A, et al. Nematode community responses to a moisture gradient and grazing along a restored riparian corridor[J]. European Journal of Soil Biology, 2012, 50: 32-38. DOI:10.1016/j.ejsobi.2011.11.006 |
| [5] |
杨锐, 吴鹏飞, 魏雪. 天然高寒草地转变为燕麦人工草地对土壤线虫群落的影响[J]. 生态学报, 2020, 40(14): 4903-4920. YANG R, WU P F, WEI X. Effects of the transformation from natural alpine grassland to artificial oat grassland on the soil nematode communities[J]. Acta Ecologica Sinica, 2020, 40(14): 4903-4920. |
| [6] |
刘贝贝, 叶成龙, 虞丽, 等. 不同植被类型的滩涂湿地土壤线虫群落特征[J]. 应用生态学报, 2012, 23(11): 3057-3064. LIU B B, YE C L, YU L, et al. Characteristics of soil nematode communities in coastal wetlands with different vegetation types[J]. Chinese Journal of Applied Ecology, 2012, 23(11): 3057-3064. |
| [7] |
周广帆, 秦洁, 祁小旭, 等. 丹江口水源涵养区退耕还草土壤线虫群落变化特征[J]. 农业资源与环境学报, 2020, 37(3): 308-318. ZHOU G F, QIN J, QI X X, et al. Effect of restoration of cultivated land to artificial grassland on soil nematodes communities in Danjiangkou water ecological conservation function area[J]. Journal of Agricultural Resources and Environment, 2020, 37(3): 308-318. |
| [8] |
EKSCHMITT K, BAKONYI G, BONGERS M, et al. Nematode community structure as indicator of soil functioning in European grassland soils[J]. European Journal of Soil Biology, 2001, 37(4): 263-268. DOI:10.1016/S1164-5563(01)01095-0 |
| [9] |
张晓珂, 梁文举, 李琪. 我国土壤线虫生态学研究进展和展望[J]. 生物多样性, 2018, 26(10): 1060-1073. ZHANG X K, LIANG W J, LI Q. Recent progress and future directions of soil nematode ecology in China[J]. Biodiversity Science, 2018, 26(10): 1060-1073. DOI:10.17520/biods.2018082 |
| [10] |
武崎, 吴鹏飞, 王群, 等. 放牧强度对高寒草地不同类群土壤动物的群落结构和多样性的影响[J]. 中国农业科学, 2016, 49(9): 1826-1834. WU Q, WU P F, WANG Q, et al. Effects of grazing intensity on the community structure and diversity of different soil fauna in alpine meadow[J]. Scientia Agricultura Sinica, 2016, 49(9): 1826-1834. |
| [11] |
海棠, 巴德嘎日布. 放牧干扰对草甸草原羊草植株及根际土壤植物寄生线虫多样性的影响[J]. 内蒙古农业大学学报, 2012, 33(3): 83-87. HAI T, Bademagaribu. Effects of grazing disturbance on the diversities of the meadow grassland Leymus chinensis pland and foundation soil pland parasitic nematides[J]. Journal of Inner Mongolia Agricultural University, 2012, 33(3): 83-87. |
| [12] |
QI S, ZHENG H, LIN Q, et al. Effects of livestock grazing intensity on soil biota in a semiarid steppe of Inner Mongolia[J]. Plant and soil, 2011, 340(1/2): 117-126. |
| [13] |
任继周. 草业科学研究方法[M]. 北京: 中国农业出版社, 1998. REN J Z. Grass industry scientific research method[M]. Beijing: China Agriculture Press, 1998. |
| [14] |
韩东亮, 贾宏涛, 朱新萍, 等. 地表水分与放牧对巴音布鲁克天鹅湖高寒湿地有机碳密度的影响[J]. 新疆农业大学学报, 2016, 39(4): 317-323. HAN D L, JIA H T, ZHU X P, et al. Influence of hydrological gradient and grazing intensity on the ecosystem organic carbon density of Bayinbulak swan lake alpine cold wetland in Xinjiang, China[J]. Journal of Xinjiang Agricultural University, 2016, 39(4): 317-323. DOI:10.3969/j.issn.1007-8614.2016.04.012 |
| [15] |
鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国科学技术出版社, 2000. LU R K. Soil agricultural chemical analysis methods[M]. Beijing: China Science and Technology Press, 2000. |
| [16] |
BARKER K R, NUSBAUM C J, NELSON L A. Seasonal population dynamics of selected plant-parasitic nematodes as measured by three extraction procedures[J]. Journal of Nematology, 1969, 1(3): 232-239. |
| [17] |
毛小芳, 李辉信, 陈小云, 等. 土壤线虫三种分离方法效率比较[J]. 生态学杂志, 2004, 23(3): 149-151. MAO X F, LI H X, CHEN X Y, et al. Extraction efficiency of soil nematodes by different methods[J]. Chinese Journal of Ecology, 2004, 23(3): 149-151. |
| [18] |
BONGERS T. The Maturity Index: An ecological measure of environmental disturbance based on nematode species composition[J]. Oecologia, 1990, 83(1): 14-19. DOI:10.1007/BF00324627 |
| [19] |
尹文英, 胡圣豪, 沈韫芬, 等. 中国土壤动物检索图鉴[M]. 北京: 科学出版社, 1998: 51-89. YIN W Y, HU S H, SHEN Y F, et al. Pictorial keys to soil animals of China[M]. Beijing: Science Press, 1998: 51-89. |
| [20] |
BONGERS T. The nematodes of the Netherlands[J]. Wageningen: Wageningen Agricultural University, 1988. |
| [21] |
BONGERS T, BONGERS M. Functional diversity of nematodes[J]. Applied Soil Ecology, 1998, 10(3): 239-251. DOI:10.1016/S0929-1393(98)00123-1 |
| [22] |
孔云, 张婷, 李刚, 等. 华北潮土线虫群落对玉米秸秆长期还田的响应[J]. 生态环境学报, 2018, 27(4): 102-108. KONG Y, ZHANG T, LI G, et al. Effects of long - term straw returning on soil nematode community in the Fluvo-aquic soil region of north China[J]. Ecology and Environmental Sciences, 2018, 27(4): 102-108. |
| [23] |
张瑞益. 北方主要草地类型土壤线虫群落结构及多样性的比较研究[D]. 长春: 东北师范大学, 2015. ZHANG R Y. Comparison of soil nematode communities structure and diversity across the main grasslands in northern China[D]. Changchun: Northeast Normal University, 2015. |
| [24] |
胡靖, 何贵勇, 尹鑫, 等. 放牧管理对青藏高原东缘高寒草甸土壤线虫的影响[J]. 土壤学报, 2016, 53(6): 1506-1516. HU J, HE G Y, YIN X, et al. Effect of grazing on soil nematode in alpine meadow on east edge of the Tibetan Plateau and its mechanism[J]. Acta Pedologica Sinica, 2016, 53(6): 1506-1516. |
| [25] |
杜晓芳, 李英滨, 刘芳, 等. 土壤微食物网结构与生态功能[J]. 应用生态学报, 2018, 29(2): 403-411. DU X F, LI Y B, LIU F, et al. Structure and ecological functions of soil micro-food web[J]. Chinese Journal of Applied Ecolog, 2018, 29(2): 403-411. |
| [26] |
乌尼尔, 张晶, 海棠. 放牧对羊草根际土壤线虫群落及区系的影响[J]. 草地学报, 2019, 27(3): 589-595. WU N E, ZHANG J, HAI T. Effects of grazing on nematode community and fauna in rhizosphere of Leymus chinensis[J]. Acta Agrestia Sinica, 2019, 27(3): 589-595. |
| [27] |
薛会英, 罗大庆, 王鸿源, 等. 藏北高寒草甸土壤线虫群落对围封及自由放牧的响应[J]. 土壤学报, 2017, 54(2): 480-492. XUE H Y, LUO D Q, WANG H Y, et al. Effects of free grazing or enclosure on soil nematodes in alpine meadows in north Tibet, China[J]. Acta Pedologica Sinica, 2017, 54(2): 480-492. |
| [28] |
WILSON M J, KHAKOULI-DUARTE T. Nematodes as environmental indicators[M]. Wallingford: CABI Press, 2009.
|
| [29] |
陈云峰, 韩雪梅, 李钰飞, 等. 线虫区系分析指示土壤食物网结构和功能研究进展[J]. 生态学报, 2014, 28(5): 1072-1084. CHEN Y F, HAN X M, LI Y F, et al. Approach of nematode fauna analysis indicate the structure and function of soil food web[J]. Acta Ecologica Sinica, 2014, 28(5): 1072-1084. |
| [30] |
薛会英, 胡锋, 罗大庆. 藏北高寒草甸植物群落对土壤线虫群落的影响[J]. 土壤学报, 2013, 33(03): 1482-1494. XUE H Y, HU F, LUO D Q. Effects of alpine meadow plant communities on soil nematode functional structure in northern Tibet, China[J]. Acta Ecologica Sinica, 2013, 33(5): 1482-1494. |
| [31] |
PORAZINSKA D L, BARDGETT R D, BLAAUW M B, et al. Relationships at the aboveground-belowground interface: Plants, soil biota, and soil processes[J]. Ecological Monographs, 2003, 73(3): 377-395. DOI:10.1890/0012-9615(2003)073[0377:RATAIP]2.0.CO;2 |
| [32] |
刘婷, 叶成龙, 陈小云, 等. 不同有机肥源及其与化肥配施对稻田土壤线虫群落结构的影响[J]. 应用生态学报, 2013, 24(12): 3508-3516. LIU T, YE C L, CHEN X Y, et al. Effects of different organic manure sources and their combinations with chemical fertilizationon soil nematode community structure in a paddy field of east China[J]. Chinese Journal of Applied Ecology, 2013, 24(12): 3508-3516. |
| [33] |
张进良. 玉米秸秆还田对土壤中微生物群落的影响[J]. 湖北农业科学, 2013, 52(12): 2744-2746. ZHANG J L. Influence of corn straw returning on the microbial community structure in soi[J]. Hubei Agricultural Science, 2013, 52(12): 2744-2746. DOI:10.3969/j.issn.0439-8114.2013.12.005 |
| [34] |
何贵永, 孙浩智, 史小明, 等. 青藏高原高寒湿地不同季节土壤理化性质对放牧模式的响应[J]. 草业学报, 2015, 24(4): 12-20. HE G Y, SUN H Z, SHI X M, et al. Soil properties of Tibetan Plateau alpine wetland affected by grazing and season[J]. Acta Prataculturae Sinica, 2015, 24(4): 12-20. |
| [35] |
PARFITT R L, YEATES G W, ROSS D J, et al. Effect of fertilizer, herbicide and grazing management of pastures on plant and soil communities[J]. Applied Soil Ecology, 2010, 45(3): 175-186. DOI:10.1016/j.apsoil.2010.03.010 |