 2014, Vol. 31
2014, Vol. 31文章信息
- 王杰, 李刚, 修伟明, 宋晓龙, 赵建宁, 杨殿林
- WANG Jie, LI Gang, XIU Wei-ming, SONG Xiao-long, ZHAO Jian-ning, YANG Dian-lin
- 氮素和水分对贝加尔针茅草原土壤酶活性和微生物量碳氮的影响
- Effects of Nitrogen and Water on Soil Enzyme Activity and Soil Microbial Biomass in Stipa baicalensis Steppe,Inner Mongolia of North China
- 农业资源与环境学报, 2014, 31(3): 237-245
- Journal of Agricultural Resources and Environment, 2014, 31(6): 513-520
- http://dx.doi.org/10.13254/j.jare.2014.0026
-
文章历史
- 收稿日期:2014-02-13

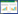




化石燃料和氮肥的大量使用,加速了全球活性氮 的固定和排放[1],大气氮沉降的增加已成为全球变化 的重要现象之一[2]。大量氮的输入(自然氮沉降和人为 施氮)不仅影响陆地生态系统可利用氮素的状况,改变 与氮相关的多种土壤生化过程[3],造成土壤富营养化, 同时还会导致土壤生态系统结构与功能的改变,进而 导致一系列环境问题的产生[4]。气候变暖与降水变化 总是相伴而生[5, 6],降水能增加土壤湿度,有助于氮肥 肥效的发挥。氮素与水分已成为影响陆地生态系统的 重要因子,因此,研究氮素与水分交互作用对陆地生 态系统土壤性质的影响具有重要的意义。
天然草地植被是地球陆地表面最大的绿色植被 层,总面积占地球陆地表面积的41%。草地生态系统 不仅具有维持生物多样性、维护全球CO2平衡和水分 循环等重要的生态功能。同时也是约占世界总人口的 17%,是总人口9.38亿人类的家园[7, 8]。草地生态系统 对陆地和大气的物质循环和能量流动至关重要[9]。随 着大气氮沉降和降水的气候变化,势必会使草地生态 系统原有的土壤结构遭到破坏,使土壤质量受到影 响。土壤酶在土壤生物化学过程中扮演重要角色,是 土壤中各种生物化学反应的催化剂,是土壤系统中最 活跃的组分之一,土壤微生物是草地生态系统的重要 组成部分,对土壤养分的供应有重要作用,在草地生 态系统的物质循环和能量流动中起到至关重要的作 用[10],其生物量对土壤条件的变化非常敏感,是草地 土壤质量变化的重要指示指标[11],大气氮沉降量的增 加和降水的变化直接影响到土壤微生物的生长和土 壤酶的活性,关系到土壤有机物的分解速率、产物形 成,进而改变土壤生态系统的碳氮循环。国内研究主 要集中于氮肥对农田生态系统的影响,本研究以贝加 尔针茅草原为研究对象,探讨氮沉降和降水变化对内 蒙古草原土壤理化性质、土壤酶活性和土壤微生物量 碳氮的变化,为深入了解我国温带草原土壤微生物对 氮沉降和降水气候变化的响应和草原合理化利用提 供理论依据和技术支撑。 1 材料与方法 1.1 试验地概况 试验地位于大兴安岭西麓,内蒙古自治区鄂温克 自治旗伊敏苏木境内,地理位置为北纬48°27′~48°35′, 东经119°35′~119°41′,海拔高度为760~770 m,地势 平坦,属于温带草甸草原区。半干旱大陆性季风气候, 年均气温-1.6益,年降水量328.7 mm,年蒸发量1 478.8 mm,≥0℃年积温2567.5℃,年均风速4 m·s-1,无霜期113 d。土壤类型为暗栗钙土[12]。植被类型为贝加尔 针茅草甸草原,建群种贝加尔针茅(Stipa baicalensis) 在群落中占绝对优势,羊草(Leymus chinensis)为优势 种,日荫菅(Carexpedif ormis)、变蒿(Artemisia commutata)、扁蓿豆(Pocockia ruthenica)、草地麻花头(Serratula yamatsutanna)、多茎野豌(Viciamul ticaulis)、祁 洲漏芦(Rhaponticu uniflorum)、寸草苔(Carex duriuscula)、肾叶唐松草(Thaictrum petaloideum)等为常见种或伴生种。共有植物66种,分属21科49属。 1.2 样地设置和土壤样品采集 1.2.1 样地设置
于2010 年6 月在围栏样地内设置养分和水分添加试验,试验采用裂区设计,主区为水分添加,副区为氮素添加处理水平。水分添加设置2 个处理,分别为不添加水分、模拟夏季增雨100 mm 灌溉,模拟增雨灌溉的时间自2010 年6 月15 日始,每7 d模拟增雨10 mm灌溉,共10 次。氮素添加处理设8个水平依次为:0(CK)、15、30、50、100、150、200、300 kg N·hm-2·a-1,分2 次施入,第1 次6 月15 日施氮50%处理水平;第2 次7 月15 日施氮50%处理水平,氮素为NH4NO3。为能够尽可能均匀施肥,根据氮处理水平,将每个小区每次所需要施加的硝酸铵(NH4NO3)溶解在8 L水中(全年增加的水量相当于新增降水1.0 mm),水溶后均匀喷施到小区内。CK 小区同时喷洒相同量的水。共16 个处理小区,6 次重复,小区面积8 m×8 m。 1.2.2 土壤样品采集
2012 年8 月,用直径为5 cm 的土钻,按照“随 机”、“等量”和“多点混合”的原则,在各个处理小区内 按照S 型取样法选取20 个点,去除表面植被,取0~ 15 cm 土壤混匀,去除根系和土壤入侵物,采用“四分 法”选取1 kg 土壤,迅速装入无菌封口袋,土样于室 内自然风干后研磨过筛,用于实验室分析。 1.3 分析方法 1.3.1 土壤理化性质的测定
土壤中的总有机碳、pH 值、全氮、硝态氮、铵态氮 和水分含量的测定分别参照鲍士旦[13]的方法。 1.3.2 土壤酶活性的测定
脲酶采用靛酚蓝比色法;磷酸酶采用磷酸苯二钠 比色法;多酚氧化酶采用乙醚萃取比色法;过氧化氢 酶采用高锰酸钾滴定法[14]。 1.3.3 土壤微生物量的测定
称取10 g过2 mm 筛的新鲜土壤,调节湿度为饱和持水量的40%,氯仿熏蒸48 h后,采用30 mL 0.5 mol·L-1K2SO4溶液浸提,用Multi N/C 3100 总有机碳/ 总氮分析仪(德国耶纳分析仪器公司)测定[15]。 1.4 数据统计与分析
采用SAS 9.1统计软件进行方差分析(ANOVA)和 相关分析(Pearson忆s correlation analysis),采用Duncan 法进行多重比较和显著性分析。 2 结果与分析 2.1 氮素和水分添加对土壤理化性质的影响
从表 1 可以看出,水分添加下,土壤pH 值呈下降趋势,且氮素添加水平越高,pH 值下降的幅度越大,高氮处理(N300)较对照处理下降4%,差异显著(P<0.05)。非水分添加处理的土壤pH值随施氮量的增加也呈下降的趋势,但是较对照处理下降幅度较小,差异不显著;相同氮素处理水平下,水分添加处理的土壤含水量普遍高于非水分添加含水量;水分添加条件下土壤有机碳含量为29.99~35.19 g·kg-1,随着氮素添加水平的增加,土壤总有机碳含量变化总体呈上升的趋势,且N50、N100、N150、N200 和N300 处理的含量显著高于其他处理(P<0.05),是对照的1.08~1.15倍。非水分添加条件下为27.53~32.46 g·kg-1,以N100和N150 处理最高,且与其他处理差异显著(P<0.05),分别是对照的1.15倍和1.16倍;不同氮添加水平的土壤全氮含量的变化趋势不同,其中,水分添加条件下,随着氮添加水平的增加,土壤全氮含量变化总体呈上升的趋势,且N50、N100、N150、N200 和N300 处理的含量显著高于其他处理(P<0.05),以N300 处理含量最高,是对照(N0)的1.12~1.28 倍。非水分添加下,随着氮添加水平的增加,全氮含量先上升后下将,均以N100 和N150 处理最高,且与其他处理差异显著(P<0.05),分别是对照的1.09 倍和1.16 倍;土壤铵态氮含量随氮肥施用量的增加而明显提高,其中水分添加条件下N200、N300 处理显著高于对照(N0)(P<0.05),分别是对照的2.19倍和4.01 倍。非水分添加条件下铵态氮含量呈先上升后下降的趋势,其中N50、N100 处理最高,分别是对照的1.98倍和2.56 倍(P<0.05);不同氮处理水平硝态氮含量变化范围较大,其中水分添加条件下,土壤硝态氮含量呈明显上升的趋势,其中N150、N200、N300 处理的硝态氮含量显著高于其他处理,且各自之间差异显著(P<0.05),是对照的7.61~33.97 倍。非水分添加条件下N100、N200 和N300 处理的硝态氮含量显著高于其他处理,且以N100处理含量最高,是对照的13.57~24.84 倍。
脲酶是一种酰胺酶,主要参与有机物分子中肽键的水解,脲酶可直接参与尿素形态转化水解生成氨、二氧化碳和水。其活性通常与土壤微生物数量、土壤有机质、全氮和速效氮等因素有关,可用来表征土壤氮素供应的强度。图 1 可以看出,2 种水分添加机制下,土壤脲酶活性随施氮水平的增加总体呈先上升后下降的趋势,其中水分添加条件下以N100 处理活性最高,显著高于对照(P<0.05)。非水分添加条件下以N30和N50处理活性最高,也显著高于对照(P<0.05)。相同氮素处理水平,水分添加条件下脲酶活性明显高于非水分添加条件下的脲酶活性,这是水分添加促进氮素有效性的主要原因。

|
| 图 1 不同氮素和水分添加处理条件下土壤脲酶活性 Figure 1 The activity of soil urease along with different N and water addition treatments |
土壤磷酸酶主要参与土壤有机磷化合物的水解, 磷酸酶的活性大小直接影响土壤有机磷的分解转化, 进而影响土壤磷素的有效性。图 2 可以看出,水分添 加条件下,随施氮水平的增加,磷酸酶活性呈上升趋 势,但较对照增加不明显。非水分添加条件下,以 N100处理活性最高,较对照差异显著。另外,除N100 处理,水分添加条件下的磷酸酶活性普遍高于非水分 添加条件下的磷酸酶活性。

|
| 图 2 不同氮素和水分添加处理条件下土壤磷酸酶活性 Figure 2 The activity of soil phosphatase along with different N and water addition treatments |
过氧化氢酶在土壤中分布较为广泛,主要参与生 物呼吸和有机物氧化过程中的物质代谢,解除氧化反 应过程中产生的过氧化氢的毒害作用,缓解生物氧化 作用对土壤和生物体造成的破坏,过氧化氢酶活性的 大小还可以表示土壤氧化过程的强度。图 3 可以看 出,水分添加条件下,随着氮处理水平的增加,过氧化氢酶活性呈上升趋势,但差异不显著。非水分添加条 件下,随着氮处理水平的增加,过氧化氢酶活性先上 升后下降,N100 处理的活性最高,且较对照差异显 著。2种水分添加机制,相同氮素水平过氧化氢酶活性 无显著差异。

|
| 图 3 不同氮素和水分添加处理条件下土壤过氧化氢酶活性 Figure 3 The activity of soil catalase along with different N and water addition treatments |
土壤多酚氧化酶主要是酶促土壤中酚类物质氧 化成醌,在土壤芳香族有机化合物转化成腐殖质的 过程中起到重要作用,在一定程度上可以表征土壤 腐殖化的程度。图 4 可以看出,水分添加条件下,随 施氮水平的增加多酚氧化酶的活性呈先上升后下降 的趋势,其中,N30 和N50 处理多酚氧化酶活性显著 高于N300 处理的。非水分添加条件下,多酚氧化酶 活性呈下降趋势,对照(N0)处理多酚氧化酶活性显 著高于N300 处理的。相同氮素水平,水分添加条件 下的多酚氧化酶活性高于非水分添加下的多酚氧化 酶活性。

|
| 图 4 不同氮素和水分添加处理条件下土壤多酚氧化酶活性 Figure 4 The activity of soil polyphenol oxidase along with different N and water addition treatments |
由图 5可见,水分添加条件下,不同氮素添加水平 的土壤微生物量碳的含量为497.04~694.79 mg·kg-1, 非水分添加条件下为334.50~552.22 mg·kg-1,低氮添 加处理(N15、N30)能增加土壤微生物量碳的含量,而 后随着氮素添加水平的增加,呈降低趋势,且氮素添 加量越大,降低的幅度越大,其中高氮处理N150、N200、N300 较对照显著降低了微生物量碳的含量 (P<0.05),水分添加与非水分添加处理下的微生物 量碳降低幅度分别为11.79%~17.55%和18.92%~ 31.64%;水分添加条件下,不同氮处理的土壤微生物 量氮的含量为58.12~74.64 mg·kg-1,非水分添加条件 下为36.69~52.05 mg·kg-1,微生物量氮含量随施氮量 的增加而增加,但是较对照变化差异不显著。另外,相 同施氮水平下,水分添加处理微生物量碳和微生物量 氮含量均高于非水分添加处理的,说明水分添加有利 于微生物的生长,而高氮肥不利于微生物生长。

|
| 图 5 不同处理土壤微生物量碳氮测定结果 Figure 5 Soil microbial biomass C and N under different treatments |
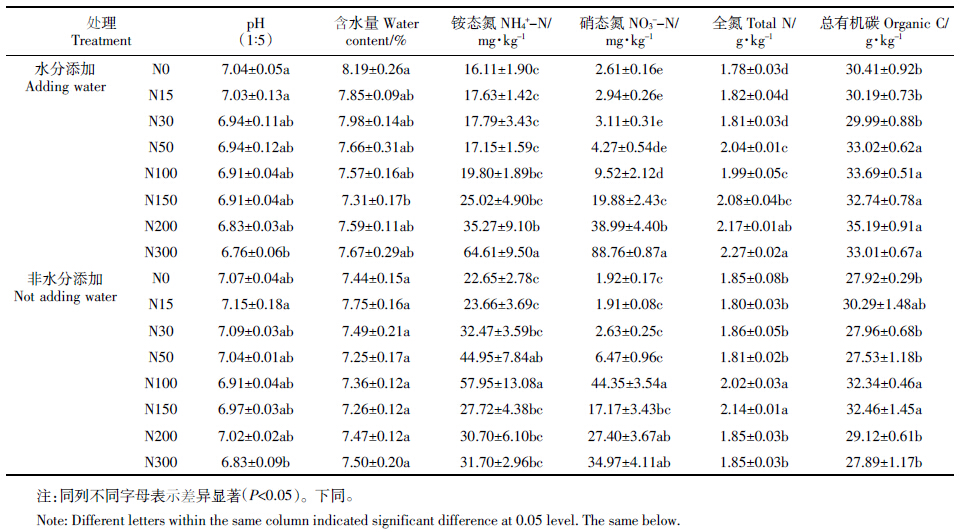
对土壤养分、酶活性做相关分析,结果表明(表 2): 多酚氧化酶活性与全氮含量呈显著负相关(P<0.05), 与铵态氮、硝态氮含量呈极显著负相关(P<0.01)。过 氧化氢酶活性与全氮和总有机碳含量呈极显著正相 关(P<0.01),与硝态氮含量呈显著正相关(P<0.05)。 说明贝加尔针茅草原土壤酶活性的高低受到土壤有 机质、氮等养分含量的制约。
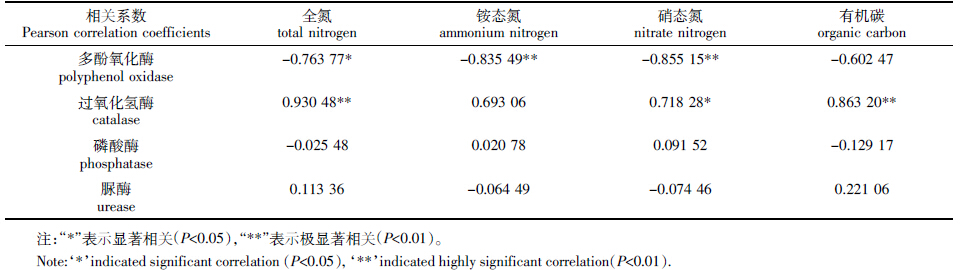 |
土壤微生物量碳氮与酶活性相关分析表明(表 3):土壤微生物量碳与多酚氧化酶活性存在极显著的 正相关关系(P<0.01),与过氧化氢酶活性存在显著负 相关关系(PP<0.05);土壤微生物量氮与过氧化氢酶呈 显著正相关(P<0.05),与多酚氧化酶呈现显著负相关 (P<0.05),与磷酸酶呈显著正相关(P<0.05)。
 |
大气氮沉降量的增加和降水格局的变化作为全球变化的主要现象之一,对草地生态系统碳氮等物质循环产生重要影响[16],通过模拟氮沉降和降水的气候变化得出,不同的氮素和水分添加处理下土壤理化性质、土壤酶活性和土壤微生物量均发生了相应的变化。长期添加氮素,土壤pH值发生了显著变化,低量氮素施入土壤后,土壤能够通过自身的缓冲能力调节pH值,使其保持稳定[17],但高量的氮素使土壤中NH4+态氮增多,硝化作用的底物浓度加大速率升高,引起土壤酸化,使土壤pH值降低[18]。曾路生等[19]的研究也表明,大量的施肥能够导致土壤的酸化,合理的肥料施用量对保持土壤环境具有重要意义,本研究中pH值与氮素添加量呈显著负相关,与铵态氮、硝态氮量呈极显著负相关(P<0.01)也能证明这一点。水分与肥料有利于地上植物的生长,凋落物和土壤植物根茬含量的增加,同时也有利于土壤微生物的生长,促进土壤腐殖质的分解,使土壤有机碳含量增加[20];水分的添加能增加土壤含水量,改善土壤物理结构,提高土壤养分的有效性,有利于铵态氮、硝态氮在土壤中的转移,本研究中水分添加条件下,高量的氮素施入,使NH4+与NO3-含量在土壤中达到超饱和状态,造成土壤盐渍化,超出植物的吸收能力,造成土壤离子累积[21]。非水分添加条件下,土壤养分含量只是在一定的范围内随着氮素添加量的增加而增加,一般N100 和N150处理的各项土壤理化指标值最高,而后下降,说明氮素的添加对植物群落氮素吸收的调控、土壤理化状况的影响存在一个阈值,超过这个阈值增加不明显或起抑制作用。
适当的氮素、水分添加处理可以提高土壤脲酶的活性,水分的添加可以增加氮肥的有效性,本研究中,水分添加条件下,土壤脲酶活性在N100 处理达到最高,非水分添加条件下以N30 和N50 处理活性最高,水分使土壤中溶解的土壤有机质的浓度升高,微生物活性增加,因而脲酶活性也随之增加[22],高氮处理使土壤中氮素含量过高,脲酶活性反而下降,徐福利等[23]的研究也表明,少量施肥,有利于地上植物的生长、根系分泌物的增加,因而土壤微生物的活性增强,土壤脲酶活性升高,但如果氮肥用量超过最大临界范围,脲酶活性将会降低;氮素添加对土壤过氧化氢酶活性影响不明显,过氧化氢酶是一类氧化还原酶,本研究中,过氧化氢酶活性与土壤全氮、有机碳呈极显著正相关,与硝态氮呈显著正相关,土壤中水分与高量有机碳、全氮含量造成了相对较强的还原状况;土壤多酚氧化酶能将土壤中的酚类物质氧化,形成腐殖质的大分子化合物,当施入氮肥时,土壤微生物和地上植物可直接利用的养分含量增加,不需要多酚氧化酶过多合成腐殖质化合物,再分解为可直接利用的养分,所以微生物和植物根系分泌量减少,酶活性降低[24]。Deforest 等[25]的研究也指出,土壤多酚氧化酶活性随着可利用性氮的增加呈下降趋势,土壤中高浓度的NO3-、NH4+抑制了分泌多酚氧化酶的真菌活性[10],本研究中,多酚氧化酶活性也与铵态氮、硝态氮含量呈极显著负相关。土壤酶是一个对外界环境变化较敏感的指标,其活性的高低主要受到土壤有机质、氮、磷等养分含量的制约,所以,土壤酶与土壤养分因子之间的相互作用机制共同影响着土壤有机质的转化过程、转化效率和土壤养分的演化方向。一方面,土壤有机质与全量养分的增加为微生物提供了充足的能源物质,促进微生物的快速繁殖和代谢过程,分泌的土壤酶数量与活性不断增强;另一方面,土壤酶活性的增强又促进了土壤有机质与全量养分的分解、速效养分的释放[26],有利于地上植物的快速生长。土壤养分、酶与微生物是土壤生态系统重要的组成部分,其中养分与酶活性对微生物的生长有重大影响,对保持土壤生态系统结构和功能稳定性起到重要作用。
高量氮素添加处理(N100~N300)能显著降低微生物量碳含量,而低量氮素添加处理能明显提高其含量,金淑兰等[27]的研究也表明,单施低浓度氮肥可以增加土壤微生物量碳,而高浓度的氮肥能明显导致土壤微生物量碳的下降,高浓度的氮肥抑制了微生物的生长和活性,产生的NO3-对微生物有直接的毒害作用,长期添加氮肥又造成了土壤质量下降,从而导致微生物活性下降[28],Andersson 等[29]研究也发现凋落物层的土壤微生物量碳随氮沉降量的增加而降低,而施用适量无机氮肥,土壤有效氮含量增加,提高了植物生产力,增加了凋落物、植物根茬等的残留,使土壤中积累的有机碳总量增加,而有机碳含量高的土壤能提供更多的有效碳源,有利于保持相对高的微生物活性[30]。Dalmonech 等[31]的研究也表明土壤微生物的活性与土壤有机碳的含量相关。而微生物量氮含量随施氮梯度的升高而增加,相关分析表明,水分添加下微生物量氮含量与土壤全氮含量呈显著正相关(P<0.05),与铵态氮和硝态氮含量呈极显著正相关(P<0.01),氮素的添加可能促进了某些喜氮微生物的生长,改变土壤微生物的群落组成[32],赵俊晔等[33]的研究也表明,施用氮肥能明显提高0~40 cm 土层的微生物量氮含量,当施氮量小于240 kg·hm-2时,微生物量氮含量随施氮量的增加而增加,氮肥的施用,促进了土壤微生物对氮素的固持,微生物量碳氮含量也表现为水分添加下较高,说明水分有利于氮肥肥效的发挥,能提高土壤微生物的活性,是影响微生物生长的重要因素,Ceder原lunda 等[34]的研究表明,含水量高的土壤有较高的微生物量与微生物活性。有研究发现土壤湿度在调节微生物活性与多样性方面至关重要,它直接影响微生物的生理状态,限制微生物分解某些化合物的能力,同时调节土壤酶和土壤理化性质,进而影响微生物组成与活性[35]。相关分析表明,土壤微生物量碳氮与土壤过氧化氢酶、多酚氧化酶和磷酸酶有显著的相关性,有研究表明,土壤过氧化氢酶和磷酸酶活性的增加能提高土壤微生物量氮含量,而多酚氧化酶活性的提高能为土壤微生物提供更多的能源物质,有利于微生物量碳的增加。土壤养分、酶与微生物是土壤生态系统重要的组成部分,三者之间相互影响、相互作用,一方面,土壤中的营养物质能为微生物生长和繁殖提供能量;另一方面,这些营养物质的循环也要依赖于微生物和酶对底物的转化和生成。在草地生态系统中,土壤的稳定性是维持草地生态系统结构和功能的稳定重要因素,是草地生态系统恢复的前提[36]。 4 结论
长期氮素和水分添加能够显著改变土壤的理化 性状,影响土壤酶活性和微生物的生长,高氮处理 (N150~N300)较显著降低了微生物量碳的含量,微生 物量氮的含量随施氮量的增加而升高。土壤理化性 质、酶活性和土壤微生物量之间具有显著相关性,三 者之间相互影响、相互作用。本研究中适量的氮素和 水分添加能改善土壤理化和生物性状,利于土壤生态 系统的恢复与发展。反之,过量的氮素添加不利于维 持草地土壤生态系统的稳定。
| [1] | 张璐,黄建辉,白永飞,等. 氮素添加对内蒙古羊草草-净氮矿化 的影响[J]. 植物生态学报,2009,33(3): 563-569. ZHANG Lu, HUANG Jian-hui, BAI Yong-fei, et al. Effects of nitrogen addition on net nitrogen mineralization in Leymus Chinensis Grassland, Inner Mongolia, China[J]. Journal of Plant Ecology, 2009, 33(3): 563-569.(in Chinese) |
| [2] | Zhou X B, Zhang Y M, Downing A. Non-linear response of microbial ac-tivity across a gradient of nitrogen addition to a soil from the Gurban-tunggut Desert, northwestern China[J]. Soil Biology and Biochemistry, 2012, 47: 67-77. |
| [3] | 张乃莉,郭继勋,王晓宇,等. 土壤微生物对气候变暖和大气 N 沉降 的响应[J]. 植物生态学报,2007,31(2): 252-261. ZHANG Nai-li, GUO Ji-xun, WANG Xiao-yu, et al. Soil microbial feedbacks to climate warming and atmospherican deposion[J]. Journal of Plant Ecolog, 2007, 31(2): 252-261.(in Chinese) |
| [4] | Xu Y B, Cai Z C. Denitrification characteristics of subtropical soils in China affected by soil parent material and land use[J]. Eur J Soil Sci, 2007, 58(6): 1293-1303. |
| [5] | IPCC. Climate change 2007: the physical science basis: summary for policymakers[R]. IPCC WGI Fourth Assessment Report, 2007. |
| [6] | 王冀,娄德君,曲金华,等. IPCC-AR4 模式资料对东北地区气候 及可利用水资源的预估研究[J]. 自然资源学报,2009,24(9): 1647-1657. WANG Ji, LOU De-jun, QU Jin-hua, et al. Prediction of climate and u-tilizable precipitation change in northeast China by using IPCC-AR4 model data[J]. Journal of Natural Resources, 2009, 24(9): 1647-1657.(in Chinese) |
| [7] | 联合国开发计划署,联合国环境规划署,世界银行,世界资源研究 所. 世界资源报告(2000—2001). 北京: 中国环境出版社,2000: 44. The United Nations Development Program(UNDP), United Nations En-vironment Programme(UNEP), the World Bank, the World Resources Institute. World resources report(2000—2001) [R]. Beijing: China En-vironmental Press, 2000: 44.(in Chinese) |
| [8] | White R S, Murray M, Rohweder. Pilot analysis of global ecosystems: grassland ecosystems technical report[R]. Washington, D C: World Re-sources Institute, 2000. |
| [9] | 赵帅,张静妮,赖 欣,等. 放牧与围栏内蒙古针茅草-土壤微生 物生物量碳、氮变化及微生物群落结构 PLFA 分析[J]. 农业环境科 学学报,2011,30(6): 1126-1134. ZHAO Shuai, ZHANG Jing-ni, LAI Xin, et al. Analysis of microbial biomass C, N and soil microbial community structure of Stipa Steppes using PLFA at grazing and fenced in Inner Mongolia, China[J]. Journal of Agro-Environment Science, 2011, 30(6): 1126-1134.(in Chinese) |
| [10] | 赵玉涛,李雪峰,韩士杰,等. 不同氮沉降水平下两种林型的主要 土壤酶活性[J]. 应用生态学报,2008,19(12): 2769-2773. ZHAO Yu-tao, LI Xue-feng, HAN Shi-jie, et al. Soil enzyme activities under two forest types as affected by different levels of nitrogen deposi-tion[J]. Chinese Journal of Applied Ecology, 2008, 19(12): 2769-2773.(in Chinese) |
| [11] | 毕明丽,宇万太,姜子绍,等. 施肥和土壤管理对土壤微生物生物 量碳、氮和群落结构的影响[J]. 生态学报,2010,30(1): 32-42. BI Ming-li, YU Wan-tai, JIANG Zi-shao, et al. Effects of fertilization and soil management on microbial biomass and community[J]. Acta E-cologicaSinica, 2010, 30(1): 32-42.(in Chinese) |
| [12] | 杨殿林,韩国栋,胡跃高,等. 放牧对贝加尔针茅草-群落植物多 样性和生产力的影响[J]. 生态学杂志,2006,25(12): 1470-1475. YANG Dian-lin, HAN Guo-dong, HU Yue-gao, et al. Effects of graz-ing intensity on plant diversity and aboveground biomass of Stipa baicalensis grassland[J]. Chinese Journal of Ecology, 2006, 25(12): 1470-1475.(in Chinese) |
| [13] | 鲍士旦. 土壤农化分析[M]援 北京: 中国农业出版社,2000: 30-83. BAO Shi-dan. Soil chemical analysis[M]. Beijing: China Agriculture Press, 2000: 30-83.(in Chinese) |
| [14] | 关松荫. 土壤酶及其研究法[M]. 北京: 农业出版社,1986: 296-339. GUAN Song-yin. Soil enzyme and its research methods[M ]. Beijing: Agriculture Press, 1986: 296-339.(in Chinese) |
| [15] | 张海芳,李刚,宋晓龙,等. 内蒙古贝加尔针茅草-不同利用方 式土壤微生物功能多样性变化[J]. 生态学杂志,2012,31(5): 1143-1149. ZHANG Hai-fang, LI Gang, SONG Xiao-long, et al. Functional diver-sity of soil microbial communities in Stipa baicalensis steppe in Inner Mongolia as affected by different land use patterns[J]. Chinese Journal of Ecology, 2012, 31(5):1143-1149.(in Chinese) |
| [16] | Zhang Y, Zheng L, Liu X, et al. Evidence for organic N deposition and its anthropogenic sources in China[J]. Atmospheric Environment, 2008, 42(5): 1035-1041. |
| [17] | Glaser K, Hackl E, Inselsbacher E, et al. Dynamics of ammonia-oxi-dizing communities in barley-planted bulk soil and rhizosphere follow-ing nitrate and ammonium fertilizer amendment[J]. FEMS Microbiology Ecology, 2010, 74: 575-591. |
| [18] | Khalil M I, Rahman M S, Schmidhalter U, et al. Nitrogen fertilizer-in-duced mineralization of soil organic C and N in six contrasting soils of Bangladesh[J]. Journal of Plant Nutrition and Soil Science, 2007, 170: 210-218. |
| [19] | 曾路生,高岩,李俊良,等. 寿光大棚菜地土壤酸化与土壤养分 变化关系研究[J]. 水土保持学报,2010,24(4): 157-161. ZENG Lu-sheng, GAO Yan, LI Jun-liang, et al. Changes of acidifica-tion and nutrient accumulation in greenhouse vegetable soils in Shouguang[J].JournalofSoilandWaterConservation,2010,24(4): 157-161.(in Chinese) |
| [20] | 曾骏,郭天文,包兴国,等. 长期施肥对土壤有机碳和无机碳的 影响[J]. 中国土壤与肥料,2008(2): 11-14. ZENG Jun, GUO Tian-wen, BAO Xing-guo, et al. Effections of soil or-ganic carbon and soil inorganic carbon under long-term fertilization[J]. Soil and Fertilizer Sciences in China, 2008(2): 11-14.(in Chinese) |
| [21] | 张英俊,杨高文,刘楠,等.草-碳汇管理对策[J].草业学报,2013,22(2): 290-299. ZHANG Ying-jun, YANG Gao-wen, LIU Nan, et al. Review of grass-land management practices for carbon sequestration [J].Acta Prata-culturae Sinica, 2013, 22(2): 290-299.(in Chinese) |
| [22] | 金继运,刘荣乐译. 土壤肥力与肥料[M]. 北京: 中国农业科技出版 社,1998: 95-155. JIN Ji-yun, LIU Rong-le. Soil fertility and fertilizer[M]. Beijing: Chi-na Agricultural Science and Technology Press, 1998: 95-155.(in Chi-nese) |
| [23] | 徐福利,梁银丽,张成娥,等. 施肥对日光温室黄瓜生长和土壤生 物学特性的影响[J]. 应用生态学报,2004,15(7): 1227-1230. XU Fu-li, LIANG Yin-li, ZHANG Cheng-e, et al. Effect of fertiliza-tion on cucumber growth and soil biological characteristics in sunlight greenhouse[J]. Chinese Journal of Applied Ecology, 2004, 15(7): 1227-1230.(in Chinese) |
| [24] | 刘磊,谷洁,高华,等. 不同施肥水平对小麦生长期土壤氧 化还-酶活性的影响[J]. 水土保持通报,2010,30(6): 13-16. LIU Lei, GU Jie, GAO Hua, et al. Effect of different fertilizer levels on soil oxidoreductases activities in wheat growing stage[J]. Bulletin of Soil and Water Conservation, 2010, 30(6): 13-16.(in Chinese) |
| [25] | Deforest J L, Zak D R, Pregitzerc K S, et al. Atmospheric nitrate depo-sition and the microbial degradation of cellobiose and vanillin in a northern hardwood forest[J]. Soil Biology and Biochemistry, 2004, 36: 965-971. |
| [26] | 蔡晓布,彭岳林,薛会英,等. 不同培肥方式对西藏中部退化土壤 微生物的影响研究[J]. 中国生态农业学报,2004,12(1): 108-111. CAI Xiao-bu, PENG Yue-lin, XUE Hui-ying, et al. Effects of different methods of applying fertilizers on the living beings in soil in central Ti-bet[J]. Chinese Journal of Eco-Agriculture, 2004, 12(1): 108-111.(in Chinese) |
| [27] | 金兰淑,郑 佳,徐 慧,等. 施氮及灌溉方式对玉米地土壤硝化 潜势及微生物量碳的影响[J]. 水土保持学报,2009,23(4): 218-226. JIN Lan-shu, ZHENG Jia, XU Hui, et al. Effect of different nitrogen condition for soil microbial biomass carbon and potential nitrification rate of corn[J]. Journal of Soil and Water Conservation, 2009, 23(4): 218-226.(in Chinese) |
| [28] | 张成霞,南志标.土壤微生物生物量的研究进展[J].草业科学,2010,27(6): 50-57. ZHANG Cheng-xia, NAN Zhi-biao. Research progress of soil micro-bial biomass in China[J]. Prataculturl Science, 2010, 27(6): 50-57.(in Chinese) |
| [29] | Andersson M, Kjφller A, Struwe S. Microbial enzyme activities in leaf litter, humus and mineral soil layers of European forests[J]. Soil Biology and Biochemistry, 2004, 36: 1527-1537. |
| [30] | Vanegas J, Landazabal G, Melgarejo L M, et al. Structural and function-al characterization of the microbial communities associated with the u-pland and irrigated rice rhizospheres in a neotropical Colombian savan-nah[J]. European Journal of Soil Biology, 2013, 55: 1-8. |
| [31] | Dalmonech D, Lagomarsino A, Moscatelli M C, et al. Microbial perfor-mance under increasing nitrogen availability in a Mediterranean forest soil[J]. Soil Biology and Biochemistry, 2010, 42: 1596-1606. |
| [32] | 罗希茜,郝晓晖,陈涛,等. 长期不同施肥对稻田土壤微生物群 落功能多样性的影响[J]. 生态学报,2009,29(2): 740-748. LUO Xi-qian, HAO Xiao-hui, CHEN Tao, et al. Effects of long-term different fertilization on microbial community functional diversity in paddy soil[J]. Acta Ecologica Sinica, 2009, 29(2): 740-748.(in Chi-nese) |
| [33] | 赵俊晔,于振文,李延奇,等. 施氮量对土壤无机氮分布和微生物 量氮含量及小麦产量的影响[J]. 植物营养与肥料学报,2006,12(4): 466-472. ZHAO Jun-ye, YU Zhen-wen, LI Yan-qi, et al. Effects of nitrogen ap-plication rate on soil inorganic nitrogen distribution, microbial biomass nitrogen content and yield of wheat[J]. |